4 Прокариоттардың морфологиясы және ультрақұрылымдық ұйымы
4.1 Прокариоттардың морфологиялық әртүрлілігі
Өсімдік табиғатындағы және құрамында хлорофилл жоқ ең кішкентай бір жасушалы организмдер бактерияларға жатады. Олар биологиялық ерекшеліктері мен қарапайым көлденең бөліну арқылы көбею қабілеті үшін фрагментті саңырауқұлақтар деп аталды. Олар Schizomycetes класына Eubacteriales ретіне жатады.
Бактериялардың мөлшері микрометрдің оннан бір бөлігінен (1 мкм=0,001 мм) ұзындығы 10–15 мкм-ге дейін және диаметрі 0,2-ден 1,0 мкм-ге дейін. Сондай-ақ айтарлықтай үлкен және кішірек организмдер бар. Тұрақты жағдайларда бактериялардың мөлшері мен формалары осы түрге, тұқымға немесе отбасына тән тұрақты болуы мүмкін, бұл олардың ұзақ эволюциясы процесінің нәтижесі. Бірақ басқа жағдайларда, мысалы, температура, осмостық қысым немесе антибиотиктер сияқты өмір сүру жағдайларының әсерінен бактериялардың мөлшері мен формасы өзгеруі мүмкін. Бактериялардың мөлшері микрометрмен (мкм) көрсетіледі. Бір микрометр 1000 нм (нанометр). Сонымен: 1 мм = 103 мкм = 106 нм = 10 А = 109 пм (пикометрлер). Бактериялардың жеке компоненттері нанометрмен де өлшенеді. Микроорганизмдердің құрылымы бүкіл жасушалардың электронды микроскопиясы және олардың ультра жұқа тілімдері арқылы зерттеледі.
Бактериялық жасуша белгілі бір өмірлік функцияны орындайтын морфологиялық және биохимиялық оқшауланған құрылымдарды қамтитын күрделі ішкі ұйымға тән тірі организм.
Бактериялық жасушалардың пішіні, мөлшері, орналасуы және жасушалардың тіркесімі сияқты морфологиялық белгілеріне сәйкес бактериялардың жіктелуі жүзеге асырылады.
Бактериялар формасы бойынша үш негізгі топқа бөлінеді:
1) сфералық (коккалар);
2) таяқша тәрізді (бактериялар, бациллалар, клостридиялар);
3) бұралған (вибриондар, спириллдер және спирохеттер).
Коккалар (Coccus) бұл бактериялар диаметрі 1–2 мкм болатын сфералық пішінді. Коккалар және басқалары бар, мысалы, бұршақ тәрізді, сфералық, ланцет тәрізді. Коккалардың бөліну бағытына байланысты ерекше формалар қалыптасады. Босануға жіктеудің негізі – коккалардың мәдениетте орналасуы. Коккалардың бір жазықтықта бөлінуімен және еншілес жасушалардың тез бөлінуімен моно немесе микрококктар (Micrococcus тұқымдасы) түзіледі. Егер бір жазықтықта бөліну кезінде еншілес жасушалар жұптасып қалса, онда диплококктар (Diplococcus тұқымдасы) немесе бірнеше біріктірілген жасушалардан тұратын тізбектер – стрептококктар (Streptococcus тұқымдасы) түзіледі. Екі өзара перпендикуляр жазықтықта пайда болатын бөліну, онда коккалар 4-ке қосылады жасушалар түзеді – тетракокктар (Tetracoccus тұқымдасы). Жасушаның өзара перпендикуляр үш бағытта бөлінуі кезінде коккалар 8,16 немесе одан да көп жасушалардан тұратын пакеттер немесе бумалар пішінін құрайды, сарциналар (тұқым Sarcina).
Коккалардың кездейсоқ бөлінуімен жүзім шоғырына ұқсас үйінділер түзіліп, стафилококктар (Staphylococcus тұқымдасы) деп аталатын формалар пайда болады (3-сурет). Микроорганизмдердің бұл тобында адамдар мен жануарлардың ауруларын тудыратын түрлер бар.

1 – моно немесе микрококкалар (туыс Micrococcus), 2 – диплококктар (туыс Diplococcus), 3 – стрептококктар (туыс Streptococcus), 4 – тетракокктар (туыс Tetracoccus), 5 – сарциналар (туыс Sarcina), 6 – стафилококктар (туыс Staphylococcus)
3-сурет – Бөлінгеннен кейін жасушалардың орналасуына байланысты коккалардың туыстық қатынасы
Таяқша тәрізді микроорганизмдер кесілген, үшкір немесе дөңгелек ұштары бар цилиндр тәрізді. Пішіндер спора түзбейтін бактериялар деп аталады (Bacterium тұқымы). Тармақталған пішінді таяқша тәрізді бактериялар коринебактерияларға жатады (Corynebacterium тұқымдасы). Таяқша тәрізді бактериялар мөлшері бойынша әр түрлі болады. Оптикалық микроскопта (0,1х0,15 мкм) көрінетін және бірнеше ондаған микрон болатын бактериялар бар.серобактериялар. Таяқша тәрізді бактериялардың мөлшері Ұзындығы 1–5 мкм аралығында болады және диаметрі 0,5–1 мкм құрайды. Қолайсыз жағдайларда кейбір таяқша тәрізді бактериялардың цитоплазмасында дөңгелек немесе сопақ пішінді дене болып табылатын спора пайда болады.
Спора түзетін микроорганизмдер бациллаларға жатады (Bacillus тұқымдасы) немесе клостридиялар деп аталады (Clostridium тұқымдасы). Таяқша тәрізді бактериялар кеңістікте ретсіз, жұптасып орналасуы мүмкін (диплобактериялар, диплобактериялар). Бұл микроорганизмдердің ең көп және әр түрлі тобы, олардың арасында сапрофитті және патогенді түрлер бар.
Бұралған бактериялар оларға бір немесе бірнеше бұйралары бар бактериялар жатады. Вибрион (Vibrio тұқымдасы) а деп аталады бактерия бір толық емес бұралу үтір түрінде спираль. Спирилла (Spirillum тұқымдасы) бірнеше спираль тәрізді бұйралары бар бактерия деп аталады. Егер микробтың ұсақ бұйралары көп болса, яғни штопор тәрізді бұралған болса, онда оны спирохета деп атайды (қатар Spirochaetales) (сурет. 4). Бактериялардың морфологиясы мен ішкі құрылымы тірі және жансыз дақылдардың препараттарында зерттеледі.
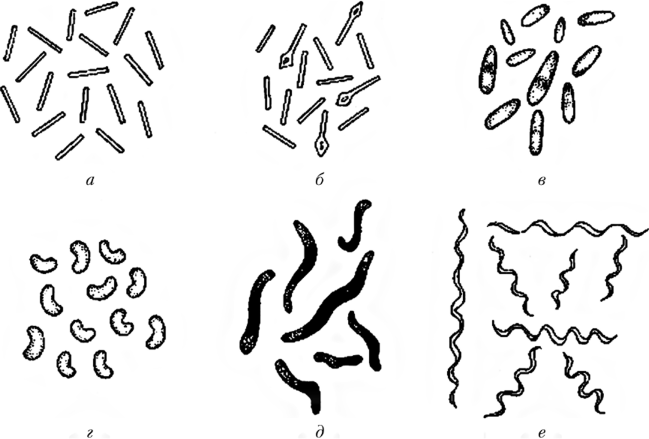
а – бактерия (туыс Bacterium), б – коринебактерия (туыс Corinebacterium), в – бацилла (туыс Bacillus), клостридия (туыс Clostridium), г – вибрион (туыс Vibrio), д - спирилла (туыс Spirillum), е - спирохетта (қатар Spirochaetales)
4-сурет – Бактериялардың таяқша тәрізді және бұралған формалары
4.2 Прокариоттардың беткі құрылымдары
Бактериялық жасуша беткі құрылымдардан мен протопласттан тұрады, беткі құрылымдарға мыналар жатады: жасуша қабырғасы, капсула, шырышты қабық, талшықтар мен түктер олар цитоплазмалық мембрананың (ЦПM) сыртында орналасқан. Протопласт бұл цитоплазмалық мембрана (ЦПM) цитоплазмамен бірге.
Жасуша қабырғасы цитоплазмалық мембрананы тығыз жабатын көп қабатты түзілім. Ол бактерия жасушасының маңызды және міндетті құрылымдық элементі болып табылады және жасушаның құрғақ затының 5–50 пайызын құрайды. Жасуша қабырғасында басқа жасуша құрылымдарында кездеспейтін арнайы полимерлі кешендер бар.
Жасуша қабырғасындағы химиялық заттардың құрамы мен құрылымы маңызды диагностикалық белгілердің бірі болып табылатын тұрақтылықпен сипатталады. Көптеген бактериялардың жасуша қабырғасында үш қабат бөлінеді: сыртқы – липопротеин, ортаңғы – липополисахарид және ішкі – мукопептид (муреин). Ішкі қабат жасуша қабырғасының қаттылығын (қаттылығын) қамтамасыз етеді. Қабырғаның құрылымына байланысты прокариоттар әр түрлі боялған және грам оң және грам теріс болып бөлінеді. Бояу әдісін 1884 жылы дат ғалымы Х. Грамм ұсынған.
Бактериялық жасушаның ішкі муреин қабатында мукополипептидтер, полифосфат нуклеотидтері және липидтер айтарлықтай мөлшерде болады. Дәл осы муреин қабаты грам оң бактерияларда айқын көрінеді. Грам теріс бактериялардағы бұл қабат шамалы болуы мүмкін. Ол жоғарыдан күшті липополисахаридті және липопротеидті қабаттармен жабылған.
Бактериялардың протопласттары, сферопласттары және L-формалары, олардың қасиеттері
Грам оң бактерияларға литикалық ферменттер, лизоцим және эндопептидазалар әсер еткенде жасуша қабырғасы жоқ жасушалар – протопластар алынады. Олар әрдайым сфералық болып табылады, оларда тыныс алу, ақуыздардың синтезі, нуклеин қышқылдары, спора түзілуі сияқты негізгі өмірлік процестер жүзеге асырылады. Протопласттардың мөлшері ұлғаюы мүмкін, азотты бекітеді және олар жасуша қабырғасын синтездей алмайды. Олар сирек бөлінеді, өйткені фаг рецепторлары жасуша қабырғасында локализацияланған протопластар фагтарды адсорбцияламайды.
Ескі дақылдарда теңгерімсіз өсу жағдайында иммундық сарысулардың әсерінен жасуша қабырғасынан ішінара айырылған бактериялық жасушалар сферопластар анықталады. Оларды гипертониялық сахароза ерітіндісінде немесе NaCl (осмостық тұрақтандырғыштар) пенициллиннің әсерінен де алуға болады. Жасуша қабырғасының ішінара сақталуын ескере отырып, сферопластар фагтарды адсорбциялауға, көбеюге қабілетті және олардың пайда болуына себеп болған факторларды жойған кезде бастапқы жасуша пішініне оңай ауыса алады. Үлкен өлшемдер, мезосома типті жасуша мембраналарының болмауы, осмостық жағдайларға жоғары сезімталдық протопласттар мен сферопласттардың жалпы қасиеттері болып табылады.
Антибиотикалық терапия кезінде немесе бактерия жасушасының қабырғасында пептидогликан биосинтезі бұзылған кезде оның ішінара немесе толық айырылуына әкеліп соқтыратын бактериялардың L-формалары пайда болады. Олардың көбею функциясы бұзылған, бірақ өсу функциясы сақталған, нәтижесінде олар алып жасушаларға дейін өседі (50 мкм дейін). Жоғары вакуолизацияланған сфералық, жіп тәрізді, алмұрт тәрізді формалар түзіледі. L-формалары метаболикалық белсенді, элементтердің бөлінуіне және бірігуіне қабілетті, пигментті орталығы мен шілтер жиегі (жұмыртқа түрінде) өсіп келе жатқан ортаға тән колониялар түрінде өседі. Патогендік бактериялардың L-формалары патогенді. L - формаларының өзгеруіне байланысты аурулар ұзақ уақыт бойы аз өліммен, бірақ мүгедектікпен жүреді. Бактериялардың осы түрлерінің пайда болуы қолайсыз жағдайларды бастан кешіру үшін бейімделу мәніне ие.
Бактерия жасушаларының жасуша қабырғасының функциялары:
1) қалыптау функциясы;
2) қоршаған орта факторларының әсерінен қорғайды;
3) ішкі осмостық қысымнан қорғайды;
4) жасушаға қажетті заттар мен иондарды тасымалдауға қатысады;
5) бұл антибиотиктер мен улы заттардың енуіне кедергі;
6) жасушаның мазмұнын гидролитикалық ферменттерден оқшаулайды;
7) құрамында тасымалдау ақуыздары мен гидролитикалық ферменттер бар;
8) құрамында арнайы рецепторлар мен антигендер бар;
9) конъюгация кезінде, сондай-ақ патогендік бактериялар мен жоғары организмдердің тіндері арасында жасушааралық өзара әрекеттесу үшін жағдай жасайды.
4.3 Шырышты беттік құрылымдар: шырышты қабаттар, капсулалар және қақпақтар Шырышты беттік құрылымдардың химиялық табиғаты мен қызметі
Көптеген прокариоттардағы жасуша қабырғасы құрылымдық ерекшеліктеріне байланысты капсула, шырышты қабат немесе қақпақ деп аталатын шырышты заттармен қоршалған. Жасуша биосинтезінде түзілген органикалық полимерлер.
Жасуша қабырғасымен байланысын сақтай отырып, жасушаны қоршап тұратын аморфты құрылымның шырышты түзілуі деп аталады капсула. Микрокапсуланың қалыңдығы 0,2 мкм-ден аз болса, ол капсула деп аталады, макрокапсулалардың қалыңдығы 0,2 мкм-ден асады. Препараттың контрастты бояуы бар жарық микроскопымен микроскопиялау кезінде макрокапсула көрінеді. Капсуланың болуына өсіру жағдайлары және бактериялардың белгілі бір штамдарына жататындығы әсер етеді. Мутация кезінде бактериялар капсула түзу қабілетін жоғалтуы мүмкін капсуласыз формалар түзеді. Капсула бактерияларды қоршаған ортаның қолайсыз әсерінен, мысалы, кептіру, сондай-ақ антиденелердің әрекеті немесе фагоцитоз сияқты ағзаның қорғаныс факторларынан қорғау құралы ретінде қызмет етеді, сонымен қатар химиялық заттардан қорғайды. Егер капсула бір-бірімен байланысқан жасушалардың бүкіл тобының айналасында пайда болса, мысалы, Azotobacter chroococcum яғни бір шырышты капсуламен қоршалған болса, оларды зооглейялар деп атайды.
Сапрофитті микробтарда капсуланың пайда болуына қоректік ортада көмірсулардың артық болуы және азотты тағамның болмауы немесе кейбір түрлер үшін төмен өсіру температурасы ықпал етеді.
Кейбір жіп тәрізді бактериялардың қақпақтары шырыштың сыртқы қабаттарының қатаюы нәтижесінде пайда болады, қақпақтар капсулалардан айырмашылығы әртүрлі құрылымы бар бірнеше қабаттарға ие. Тотықсызданған металл қосылыстарының тотығуымен бактериялардың метаболизмі кезінде тотықтар қақпақтарда тұндырылады, мысалы, темір бактерияларында темір гидроксиді тұндырылады және жасушалар ерекше күшке ие болады.
Прокариот капсулаларының көпшілігінде негізгі химиялық компонент гомо немесе гетерополисахаридтер болып табылады.
Функционалды түрде капсулалар мен қақпақтар жасушаны механикалық зақымданудан, кептіруден қорғайды, қосымша осмостық тосқауыл жасайды, фагтардың енуіне жол бермейді, қосалқы қоректік заттардың көзі бола алады, көрші жасушалар арасындағы байланысты жүзеге асырады және жасушалардың әртүрлі беттерге жабысуына ықпал етеді.
4.4 Талшықтар, пилилер (фимбриялар) түрлері, құрылымы және функцияларының әртүрлілігі
Бактериялардың кейбір түрлерінде талшық деп аталатын қозғалыс мүшелері бар. Бұл өте жұқа (0,01–0,05 мкм), ұзын бұрандалы бұралған талшықтар. Олардың саны, өлшемдері және орналасуы белгілі бір түрлер үшін тұрақты.
Флагеллалардың орналасуы бойынша қозғалмалы бактериялар төрт топқа бөлінеді: монотрихтер ұшында бір флагелла орналасқан бактериялар; лофотрихтер бір ұшында флагелла шоғыры бар бактериялық жасушалар; амфитрихтерде екі полюсте бір немесе бірнеше флагелла болады; перитрихтерде жасушаның бүкіл бетінде көптеген флагелла болады (сурет. 5).
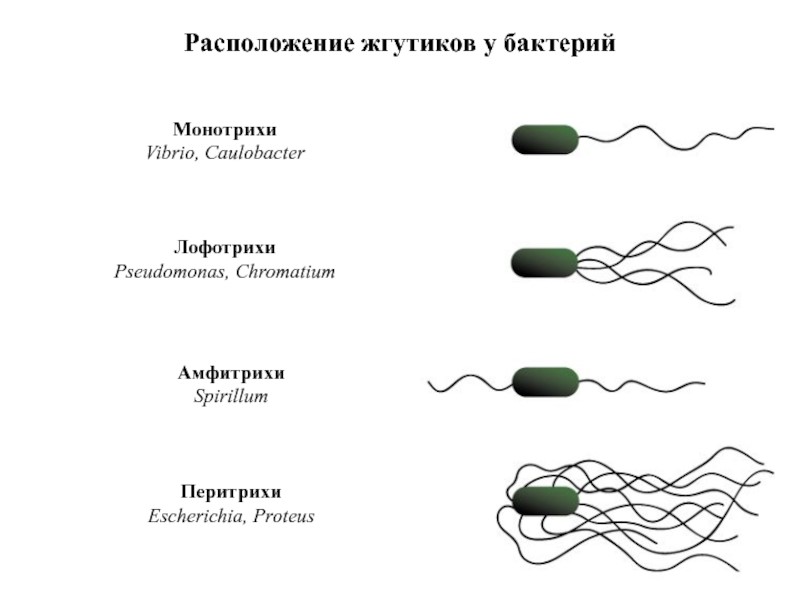
5-сурет – Бактериялардағы талшықтардың орналасуы
Талшықтар протоплазма ақуыздарынан ерекшеленетін антигендік қасиеттері бойынша фибриллярлы флагеллин ақуызынан тұрады. Морфологиялық тұрғыдан флагелла үш бөліктен тұрады бұл спиральды жіп, «Ілмек» бөлігі жасуша бетіне жақын және базальды дене. Ілмек жіптің базальды денемен икемді байланысын қамтамасыз етеді және флагелиннен ерекшеленетін ақуыздан тұрады – талшықтың фибриллярлық ақуызы.
Бактериялардың қозғалыс үлгісі фибриллалардың ритақты жиырылуына байланысты пайда болатын флагеллалардың спираль тәрізді немесе толқын тәрізді қозғалысын анықтайды. Монотрих қозғалысы әрқашан түзу сызықта жүреді, кейде бір жерде шұңқыр тәрізді қозғалыстар жасалады. Перетрихтер ретсіз құлау қозғалыстарын жасайды. Сыртқы факторлар жылжымалы бактериялардың белсенді қозғалу бағытын анықтайды. Бактериялардың химиялық зат көзіне қатысты қозғалысы химотаксис деп аталады. Осыған байланысты химиялық заттар екі топқа бөлінеді: инертті және қоздырғыш таксилер (эффекторлар). Эффекторлар тартқыштар (тартымды заттар) және репелленттер (бактерияларды тежейтін заттар) болуы мүмкін. Аттрактанттарға аминқышқылдары, дәрумендер, нуклеотидтер, қант кіруі мүмкін. Репелленттер-спирттер, фенолдар, бейорганикалық иондар, сонымен қатар аминқышқылдары және басқалар. Аэротаксис сонымен қатар химотакстың көрінісі болып табылады. Фототрофты бактерияларға тән жарыққа қарай қозғалыс деп аталады фототаксис. Бактериялардың жердің немесе магниттің магнит өрісінің сызықтары бойынша қозғалу қабілеті – магнитотаксис. Магнитотаксис жасайтын бактериялардың жасушаларында магнетит (Fe3O4) түрінде темірмен толтырылған бөлшектер табылды, олар магниттік көрсеткі қызметін атқарады және магнитосомалар деп аталады. Бактериялардың қозғалысы ерітіндінің тұтқырлығын жоғарылату немесе төмендету бағытында қозғалады деп аталады вискозитаксис.
Кірпікшелер (фимбрия, пили, ворсинкалар) бактерия жасушасында орналасқан. Қозғалысқа ешқандай қатысы жоқ және бактериялардың қозғалмалы және қозғалмайтын түрлерінде кездеседі. Бұл пилин ақуызынан тұратын, жасуша бетінен таралатын түзу, жұқа және қысқа ақуыз цилиндрлері, талшықтарға қарағанда жұқа, олардың ұзындығы 0,2–2,0 мкм, диаметрі 5–140 нм. Олар перитрихиялық типте немесе полюстерде орналасқан. Жалпы типтегі және жыныстық типтегі фимбриялардың екі түрі зерттелген. Жалпы типтегі фимбриялар бактерия жасушасына гидрофобты қасиет береді және топырақ бөлшектерінде, өсімдіктерде және жануарлар жасушаларында бактериялардың адсорбциясын қамтамасыз етеді. Жыныстық ворсинкалар немесе F-пили жыныстық процесте бактериялардың бір-біріне жабысуына ықпал етеді. F-пили конъюгациялық туннель ретінде донор жасушасынан реципиентке ДНҚ-ның берілуін қамтамасыз етеді. Фимбриялар қосымша жасушалық құрылымдарға жатады.
4.5 Цитоплазмалық мембрана, оның құрамы, құрылымы және бактериялардағы қызметі
Цитоплазмалық мембрана (ЦПM) жасушаның мазмұнын оның қабырғасынан бөледі. Бұл кез-келген жасушаның міндетті құрылымдық элементі және ЦПM тұтастығының бұзылуы өлімге әкеледі. ЦПM жасушалардың құрғақ заттарының 8–15 % құрайды, прокариот жасушаларының көпшілігінде жалғыз мембрана бар. Бактериялардың цитоплазмалық мембранасы бұл ақуыздар мен липидтерден тұратын биологиялық мембрана, кейде көмірсулардың едәуір мөлшері кездеседі. ЦПM липидтерінің қызметі механикалық тұрақтылықты сақтау және гидрофобтылықты қамтамасыз ету болып табылады
Цитоплазмалық мембрана ақуыздары ферменттер болып табылады. Аминқышқылдарының құрамы бойынша олар басқа жасушалық ақуыздардан ерекшеленеді.
Цитоплазмалық мембрана келесі функцияларды орындайды:
1) тосқауыл функциясы;
2) ЦПM-да ферменттер локализацияланған, сондықтан функциялардың бірі ферментативті болып табылады;
3) ЦПМ-да электронды тасымалдау тізбегінің тасымалдаушылары орналасқан, сондықтан ол энергетикалық функцияны орындайды;
4) хромосоманың репликациясына қатысады;
5) жасушаны біріктіреді, яғни біртұтас тұтастыққа біріктіреді;
6) тасымалдау функциясы (мембраналық тасымалдаудың әртүрлі механизмдері қолданылады).
Көлік жүйелерінің 4 түрі бар: белсенді тасымалдау, химиялық түрлендірілген молекулаларды тасымалдау, пассивті диффузия және жеңілдетілген диффузия
4.6 Бактериялардың ішкіплазмалық мембраналық құрылымдары
Цитоплазмаға терең ену және цитоплазмалық мембрананың өсуі ішкі плазмалық мембрананың пайда болуына ықпал етеді. Олардың арасында:
1) пішіні, мөлшері және жасушадағы орналасуы бойынша ерекшеленетін жергілікті ЦПМ-лар – мезосомалар деп аталады;
2) фотосинтетикалық бактерияларда фотосинтетикалық мембраналар (тилакоидтар) болады. Жасыл бактерияларда пигменттер хлоросомаларда, цианобактериялардың фикобилисомаларда кездеседі;
3) Кальвин цикліндегі СО2 фиксациясының негізгі ферменті рибулозодифосфат карбоксилазасында карбоксисомалар бар;
4) су бактерияларында, кейбір топырақ бактерияларында және балшық тұрғындарының бактерияларында газ вакуольдері немесе аэросомалар қалқымалылықты реттейді;
5) магнитосомалар – жоғарыда айтылғандай, магнитотаксисі бар бактериялардың жасушаларында магнитосомаларда Fe3O4 бөлшектері болады.
4.7 Прокариот жасушасының цитоплазмасы, рибосомалар
CPM қоршалған жасушаның ішкі құрылымы цитоплазма деп аталады. Құрамында цитозол (жартылай сұйық коллоидты масса) бар ферменттік ақуыздар, метаболикалық реакциялардың өнімдері мен субстраттары және еритін РНҚ жиынтығы. Цитоплазманың басқа бөлігіне әртүрлі құрылымдық элементтер жатады: цитоплазмалық мембраналар, рибосомалар, генетикалық аппарат және қосындылар.
Бактериялардың рибосомалары әртүрлі РНҚ молекулалары мен ақуыздардан тұратын күрделі түзілімдер. Олар ақуыз синтезделетін орын. Олардың өлшемдері 15–20 нм. Ақуыз синтезінің қарқындылығына байланысты мөлшер 5000-нан 90000-ға дейін өзгереді. Бұл 30S және 50S тұндыру коэффициенті бар екі суббөлшектен тұратын рибонуклеин бөлшектері. Прокариоттық рибосомалар 70S тұндыру константасына ие. Рибосомалық ақуыздар құрылымдық функцияны орындайды.
Рибосомалардан тұратын агрегаттар, яғни ақуыз синтезін жүзеге асыратын хабаршы және тасымалдаушы РНҚ молекулалары, полирибосомалар немесе полисомалар деп аталады. Олар мембраналық құрылымдармен байланысты болуы мүмкін немесе цитоплазмада болуы мүмкін.
4.8 Қосалқы заттар
Бактериялардың қорындағы заттар – полисахаридтер (крахмал, гранулеза, гликоген), липидтер, полипептидтер, волютин, күкірт шөгінділері.
Кейбір серобактериялардың жасушаларында әк денелері (кальций карбонаты) табылды, олар жасушаішілік күкірт тотығу кезінде пайда болатын күкірт қышқылымен қосылып, ортаны бейтараптандыру функциясын орындайды деген болжам бар.
Параспоральды қосындылар спораларға жақын кристалл тәрізді қосындылар деп аталады. Мұндай қосындыларда, мысалы, Bacillus thuringiensis грам оң топырақ бактериялары бар. Бұл қосындылар ақуыздық сипатқа ие және жәндіктер үшін улы, бірақ сонымен бірге омыртқалы жануарлар мен өсімдіктерге мүлдем зиянсыз. Бактериялардың бұл түрі ауыл шаруашылығында инсектицид ретінде кеңінен қолданылады.
Қосалқы заттар прокариоттар бұл жоғары молекулалы полимерлі молекулалар, олар кейде цитоплазмадан ақуыз мембранасымен бөлініп, осмостық белсенді емес күйде болады. Оларды осы күйде табу өте маңызды, өйткені осмотикалық белсенді заттардың көптеген молекулаларының цитоплазмасында шоғырлануы жасушаға теріс әсер етуі мүмкін.
4.9 Прокариоттардың генетикалық аппараты Бактериялардың нуклеоиды, құрылымы, мөлшері, қызметі және оны анықтау әдістері
Прокариоттардың ДНҚ-лы (нуклеоид) – бұл тығыз зат, цитоплазмада орталық аймақты алады және одан мембранамен бөлінбейді. Прокариоттарда бір ДНҚ молекуласында барлық генетикалық ақпарат бар, бактериялық хромосома деп аталады және ковалентті жабық сақина түрінде болады. Ашылған кезде молекуланың ұзындығы 1 мм-ден асуы мүмкін.
ДНҚ репликациясы жартылай консервативті механизм арқылы жүреді. Ол сақина хромосомасының цитоплазмалық мембранаға қосылу нүктесінен басталады, онда репликация процесіне жауап беретін ферментативті аппарат орналасқан
ДНҚ-ны цитоплазмалық мембранамен байланыстыру мезосомалардың қатысуымен жүзеге асырылады. Репликация процесі екі қарама-қарсы бағытта жүреді. Пайда болған еншілес хромосомалар мембранаға жабысып қалады. ДНҚ молекулаларының репликациясымен қатар ДНҚ-ның цитоплазмалық мембранамен жанасу аймағында мембрана синтезі жүреді. Нәтижесінде еншілес ДНҚ молекулалары бөлініп оқшауланған хромосомалар пайда болады
Бактериялардың жасушаларында хромосомалардан басқа ДНҚ сақинасында қамалған плазмидалар жиі кездеседі, олар тәуелсіз репликацияға қабілетті. Плазмидалар хромосомалардан ерекшеленбейді, құрамында тек белгілі бір жағдайларда қажет болатын қосымша гендер бар. Плазмидаларда антибиотиктерге төзімділік механизмдері, ерекше заттардың жойылу механизмдері және т.б. сондай-ақ плазмидаларда азотфиксацияға қажетті nif-гендер бар. Плазмидалардың еншілес жасушаларда сақталуы арнайы таралу механизмдерін қамтамасыз етеді.
Транспозондар – бактериялардың ДНҚ-сындағы қозғалмалы сегменттер, олар хромосоманың бір бөлігінен екіншісіне немесе ДНҚ хромосомадан тыс, сондай-ақ басқа жасушаларға ауыса алады. Мобильді ДНҚ сегменттері автономды репликацияға қабілетсіз.
4.10 Бактериялардың бөлінуі Бөлу тәсілдерінің әртүрлілігі
Бактериялар екі бірдей еншілес жасушалар түзілетін көлденең екілік бөлінумен сипатталады. Грам оң бөліну септа арқылы жүреді, яғни перифериядан центрге қарай созылатын көлденең септумды синтездеу арқылы, грам теріс тарылу арқылы жүреді. Бактериялардың көбеюінің басқа түрлері – бүршіктену және көптік бөліну. Бүршіктену қосымша өсу арқылы жүреді, ал аналық және аналық жасушалар бөлінеді, бүршіктенуді тең емес екілік бөліну деп санауға болады. Көптік бөліну кейбір цианобактерияларда байқалады, бұл бөліну кезінде көптеген ұсақ, кейде қозғалмалы баециттер пайда болады.
Қос бөліну бір немесе бірнеше жазықтықта жүреді. Бір жазықтықта бөлінгенде жасушалар бөлінбейді, бұл таяқша тәрізді немесе сфералық жасушалар тізбегінің пайда болуына әкеледі. Бөліну кезінде бірнеше жазықтықта әртүрлі пішіндегі жасушалық кластерлер пайда болады.
4.11 Бактериялардың дифференциациясының түрлері
Бактерия жасушасының өмір сүру деңгейі жоғары болады, сондықтан ол вегетативті жасушалардың морфологиялық дифференциациясымен сипатталады. Мамандандырылған жасушалар бактериялардың вегетативті жасушаларынан ерекшеленеді, олардың пайда болу қабілеті генетикалық аппаратта бағдарламаланған. Бактериялардың даму процесінде сыртқы факторлардың әсерінен тыныштық формалары санатына жататын құрылымдар қалыптасады. Олардың мақсаты қолайсыз жағдайларда түрдің ұзақ уақыт сақталуын қамтамасыз ету. Қолайлы жағдайлардың басталуымен немесе енуімен тыныштық формалар өніп, вегетативті жасушаларды тудырады.
Сондай-ақ көбеюге арналған морфологиялық сараланған жасушалар бар. Бұл цианобактериялардың баециттері мен гормогониялары.
Цианобактериялардың гетероцистасының арнайы метаболикалық функциялары бар морфологиялық сараланған жасушалар, түйін бактерияларының бактероидтары атмосфераның молекулалық азотын бекітеді.
Бактериялардағы споралар – бұл тыныштықтың бір түрі, оның негізгі қызметі – тұқым қуалайтын ақпаратты қолайсыз жағдайларда сақтау, саңырауқұлақ спораларынан айырмашылығы – көбею әдісі емес. Споралар сопақ, дөңгелек немесе эллипс тәрізді, механикалық әсерге төзімділікті арттыру үшін кейбір түрлерде «қаттылық қабырғалары» болады.
Аналық жасушаның цитоплазмасында пайда болатын грам оң бактериялардың тыныштықтағы жасушаларының ерекше түрі деп аталады эндоспоралар. Бір бактериялық жасуша бір эндоспораны құрайды. Эндоспораларда көп қабатты ақуыз жамылғысы, сыртқы және ішкі мембраналар және қабықтың ең қалың қабаты – кортекс бар. Кейбір бактериялар экзоспориум деп аталатын липопротеидтер мен көмірсулардан тұратын бактериялардың капсуласына ұқсас екі-үш қабатты желатинді жабынды жасайды. Эндоспоралар вегетативті жасушалар үшін өлімге әкелетін жоғары температура мен радиацияға төзімді.
Егер спора түзетін аэробты бактерияларда диаметрі жасуша диаметрінен аспайтын спора пайда болса, бактериялар бациллалар деп аталады. Спораның мөлшері жасушаның диаметрінен асатын спора түзетін анаэробты бактериялар, сондықтан олар шпиндель түрінде болады, клостридиялар деп аталады (лат. Сlostridium – ұршық).
Актиномицеттердің көпшілігінің гифалары бөлімдерді бөлімдерге бөледі, олардың әрқайсысы болашақ экзоспораны білдіреді. Экзоспоралар микробтық жасушаның сыртында түзіледі. Олардың қалыптасуы жасуша қабырғасының тығыздалуымен және қалыңдауымен қатар жүреді. Олар эндоспоралардан кортекс пен экзоспориумның болмауымен ерекшеленеді. Экзоспораларда дипиколин қышқылы жоқ. Актиномицет сопоралары бір мезгілде тыныштық жасушалары және репродуктивті құрылымдар болып табылады. Экзоспоралар Methylosinus және Rhodomicrobium тұқымдас бактериялардағы сияқты аналық жасуша полюстерінің бірінен бүршіктену арқылы пайда болуы мүмкін. Бактериялардың әртүрлі топтарында кездесетін кисталар морфологиялық тұрғыдан вегетативті жасушалардан ерекшеленбеуі мүмкін. Кисталар сонымен қатар қоршаған ортаның қолайсыз жағдайларын күту үшін тіршіліктің тыныштық түрі болып табылады. Кисталардың қабырғалары бірдей немесе әртүрлі химиялық құрамдағы бір немесе бірнеше сыртқы (экзин) және ішкі (интин) қабаттардан тұрады, сонымен қатар құрылымы әртүрлі. Цисталардың цитоплазмасында су аз, энергия мен заттардың алмасуы айтарлықтай баяулайды.
Кейбір цианобактериялардың тыныштық жасушалары акинеттер деп аталады. Микроорганизмдердің көптеген түрлерінің тыныштық жасушаларынан айырмашылығы, олар вегетативті жасушаларға қарағанда үлкенірек. Ұзын немесе сфералық, түйіршікті мазмұны және қалың қабығы бар. Олар пайда болғаннан кейін немесе жаңа өсіп келе жатқан ортаға ауысқаннан кейін бірден өніп шығуы мүмкін.
Кисталар мен акинеттер вегетативті жасушаларда физикалық әсерге төзімділік бұзылған жағдайда пайда болады. Олар вегетативті жасушаларға қарағанда төзімді, мысалы, қыздыру және кептіру.
Өзіндік жұмыс істеуге арналған тапсырмалар
- Микроорганизмдерге жататын организмдер топтары
- Прокариоттар мен эукариоттардың негізгі айырмашылығы неде?
- Бактериялардың морфологиялық және физиологиялық ерекшеліктері
- Грам оң және грам теріс бактериялардың жасуша қабырғасының ерекшеліктері қандай. Оның функциялары. Граммен бояу әдісінің мәні.
- Грам оң және грам теріс эубактериялардың жасуша қабырғаларының химиялық құрамы. Бактериялардың граммен бояуға қатынасы.
- Бактериялардың протопласттары, сферопласттары және L формалары дегеніміз не және аурудың қайталануы қалай пайда болады?
- Бактериялардың цитоплазмалық мембранасының құрылымы мен қызметі қандай? Цитоплазмалық мембрананың туындыларын атаңыз. Олардың жасушадағы қызметі қандай?
- Цитоплазма және оның қосындылары.
- Прокариоттардың сыртқы құрылымдық түзілімдері, (капсула, эндоспора, флагелла, фибрия, пили) және олардың функциялары.
- Прокариоттық микроорганизмдердің мөлшері қандай?
- Бактериялардың формалары.
- Бактерия жасушасының негізгі құрылымдарын атаңыз.
- Бактерия жасушасының қосымша құрылымдарын тізімдеңіз.
- Бактериялардың жасуша қабырғасының құрылымы мен қызметі.
- Нуклеоид дегеніміз не? Оның құрылымы мен функциялары.
- Флагелла, фимбрия және бактериялық аралар арасындағы айырмашылық неде?